
Сейчас на сайте
Сейчас на сайте 0 users и 1 guest.

Главное меню
ВЛИЯНИЕ ЗАСУХИ НА РАСТЕНИЯ ВИНОГРАДА В УСЛОВИЯХ КУЛЬТУРЫ IN VITRO
Разместил Владимир Большаков в Втр, 03/02/2010 - 14:02
ВЛИЯНИЕ ЗАСУХИ НА РАСТЕНИЯ ВИНОГРАДА В УСЛОВИЯХ КУЛЬТУРЫ IN VITRO
Е.И. Ковбасюк
Национальный научный центр
«Институт виноградарства и виноделия им. В.Е. Таирова», Украина
Проблема устойчивости сельскохозяйственных растений к неблагоприятным условиям среды была и остается актуальной. Для определения устойчивости растений выбирают различные критерии. Для винограда степень устойчивости сорта к экстремальным условиям среды, способность противостоять им, можно охарактеризовать по изменению продуктивности [3, 8]. Но для винограда этот критерий не всегда удобен, так как растение вступает в фазу плодоношения в среднем на третий год жизни, а для характеристики ценных исходных форм желательно получать такую оценку на начальных этапах работы. Поэтому нужны надежные, объективные и в то же время простые и быстрые методы. Для этого целесообразно проводить исследования с использованием культуры тканей in vitro, что ускорит и облегчит работу.
Целью наших исследований была разработка экспресс-методов для оценки засухоустойчивости винограда в условиях культуры тканей in vitro. Для этого мы использовали осмотически-активное вещество – полиэтиленгликоль (ПЭГ), который не токсичен, не попадает в растение и вызывает водный стресс у растений, уменьшая водный потенциал в питательной среде [2, 4], а также исследовали изменение в стрессовых условиях флуоресценции хлорофилла листьев винограда с помощью хронофлуориметра «Флоратест». Чтобы подтвердить действенность разработанных методов, проводили определение общепринятых физиологических показателей.
Работа выполнена в отделе питомниководства и размножения винограда ННЦ «ИВиВ им. В.Е. Таирова» на микроклонах подвойных сортов винограда Добрыня и Таировский 1. Засуху создавали путем внесения в питательную среду Мурасиге-Скуга ПЭГ концентрации 4-8%. Микроклоны контрольных вариантов культивировали на питательных средах без ПЭГ. На стрессовые среды пересаживали микроклоны с хорошо развитой корневой системой. В листьях растений проводили анализы по содержанию общей и легкоудерживаемой воды, интенсивности транспирации, содержанию пигментов [7], индукции флуоресценции хлорофилла листьев [1].
С целью изучить влияние различных концентраций ПЭГ на ростовые процессы, учитывали изменение высоты растений, количества листьев и междоузлий микроклонов в ходе исследований.
В результате исследований выявлено, что контрольные растения восстанавливали рост побега и корневой системы, тогда как на стрессовых средах уже на пятый день у микроклонов наблюдали снижение тургора листьев верхнего и среднего яруса (особенно у вариантах с содержанием ПЭГ 4-5 %), потемнение листовых пластинок, а в дальнейшем - усыхание верхушек побегов (в вариантах с содержанием ПЭГ 6-8 %). В таблице 1 показаны изменения высоты растений под действием индуцированного ПЭГ водного стресса. Так, на среде с содержанием 7,0-7,5% ПЭГ высота микроклонов винограда сорта Добрыня уменьшалась в 1,2-1,4 раза по сравнению с начальной, для сорта Таировский 1 такие изменения наблюдали уже при концентрации 6% ПЭГ в среде. За счет отмирания листьев их количество тоже уменьшалось – для сорта Добрыня в вариантах 5-6% ПЭГ в 1,1-1,2 раза и в 1,5-1,7 раз для вариантов 7-8% ПЭГ. У контрольных растений этого сорта количество листьев наоборот возрастало в 1,2 раза. Подобную тенденцию отметили для сорта Таировский 1, но изменения показателей роста, индуцированные ПЭГ, были значительнее.
Таблица 1 - БИОЛОГИЧЕСКИЕ ПОКАЗАТЕЛИ РОСТА РАСТЕНИЙ НА СРЕДЕ MС З РАЗНЫМ СОДЕРЖАНИЕМ ПЭГ
|
Сорт |
Вариант | В начале исследований | Через 10 дней |
||
|
высота растения, см | количество листьев, шт |
высота растения, см | количество листьев, шт |
||
|
Добрыня
|
Контроль 5,0% ПЭГ 6,0% ПЭГ 7,0% ПЭГ 7,5% ПЭГ 8,0% ПЭГ |
6,5 5,8 5,3 5,8 7,1 7,7 |
6,4 7,0 6,0 5,6 6,4 8,4 |
7,4 5,6 5,6 4,7 5,0 6,5 |
7,4 5,8 5,2 3,2 3,6 5,6 |
Таировский1 |
Контроль 4,0% ПЭГ 5,0% ПЭГ 6,0% ПЭГ 7,0% ПЭГ 7,5% ПЭГ 8,0% ПЭГ |
5,8 7,0 6,7 6,0 6,2 6,5 5,9 |
5,7 6,2 6,0 5,7 6,2 5,8 6,2 |
6,5 6,4 6,6 5,1 5,5 6,2 4,3 |
6,7 4,3 5,4 3,2 3,8 4,5 2, |
Во многих научных исследованиях показана тесная взаимосвязь индукционных процессов флуоресценции хлорофилла листьев с функционированием фотосинтетического аппарата и физиологическим состоянием растения в целом [1, 5]. С учетом этого мы поставили задание показать возможность использования метода индукции флуоресценции хлорофилла (ИФХ) листьев для оценки засухоустойчивости микроклонов винограда по показателям флуоресценции. Исследование ИФХ листьев растений, которые содержались на питательных средах с разным содержанием ПЭГ от 4,0 до 7,5%, показало увеличение амплитуды кривой ИФХ по сравнению с контролем. Причем наблюдается прямая зависимость кинетического уровня кривой от концентрации ПЭГ. Согласно рис. 2 на самом высоком кинетическом уровне расположены кривые ИФХ листьев растений в вариантах МС+6, 7,0 и 7,5% ПЭГ. Дальнейшее увеличение концентрации осмотического вещества до 8 % не всегда давало возможность получить четкий всплеск ИФХ и вызывало почти полную ее потерю, что вероятнее всего связано с разрушающим влиянием обезвоживания тканей на электрон-транспортную цепь и систему фотораспада воды. У растений сорта Добрыня отличия проявлялись в меньшем расхождении амплитуд кривых ИФХ, во времени выхода на стационарный уровень и длительности пребывания на новом уровне излучения. Для этого сорта выход флуоресценции на стационарный уровень наблюдали во всех вариантах, но в контроле это происходило раньше (на 71247-96458 мс) - это 62-63-я точка, а опытных вариантах – на 73-75-й точке исследований. Этот показатель свидетельствует об усиленном оттоке электронов на вторичные акцепторы и фиксации углекислого газа. У растений сорта Таировский 1 наблюдали значительные отличия между амплитудами кривых ИФХ под действием высоких концентраций ПЭГ; за 3 мин исследований выход на стационарный уровень наблюдали в 81-83-й точке.
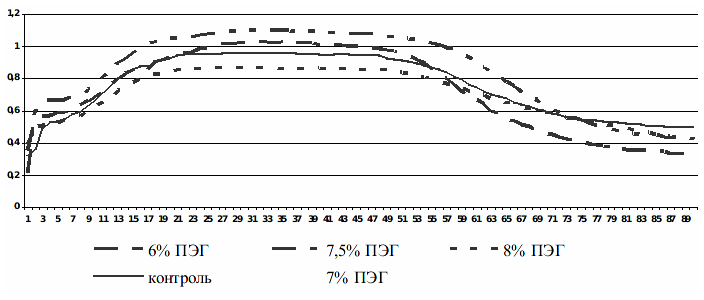
Рисунок 2 - Изменения индукции флуоресценции хлорофилла листьев винограда сорта Добрыня под влиянием модельной засухи.
Относительно биологических особенностей этих двух сортов надо заметить, что согласно нашим исследованиям, подвойный сорт Добрыня более засухоустойчив, чем Таировский 1.
Влияние модельной засухи, созданной внесением ПЭГ в питательную среду, подтвердили с помощью физиологических анализов микроклонов. Для этого исследовали количество пигментов в тканях растений. Оказалось, что растения контрольного и опытных вариантов двух сортов существенно отличались по содержанию хлорофиллов и каротиноидов. Так, содержание хлорофиллов в растениях опытных вариантов сорта Добрыня составляло 2,913 мг/г сырого веса, в вариантах с ПЭГ этот показатель был ниже в 1,2-1,8 раз, причем минимальными были значения в вариантах 6 и 7,5% ПЭГ. В микроклонах контрольного варианта определили 0,602 мг/г сырого веса каротиноидов, в опытных вариантах их количество уменьшалось до 0,372-0,544 мг/г; это в 1,1-1,6 раз ниже контрольных значений. Для сорта Таировский 1 также отметили снижение содержания пигментов в тканях. Наши результаты согласуются с данными других исследователей, которые утверждают, что под действием водного стресса в растительном организме включаются защитные механизмы адаптации [6, 8], это проявляется в уменьшении содержания хлорофиллов, чаще всего хлорофилла а, и каротиноидов.
Интенсивность транспирации и содержание легкоудерживаемой воды в тканях растений контрольных вариантов двух сортов в общем были ниже, чем в опытных (таблица 2). Количество легкоудерживаемой воды в растениях опытных вариантов было ниже по сравнению с контролем на 31,3-49,4 % (сорт Добрыня) и на 14,5-34,3% (сорт Таировский 1). Во всех вариантах с содержанием ПЭГ отмечено уменьшение количества общей воды по сравнению с контролем на 9,0-17,9% для сорта Добрыня и на 5,5-16,1% для сорта Таировский 1.
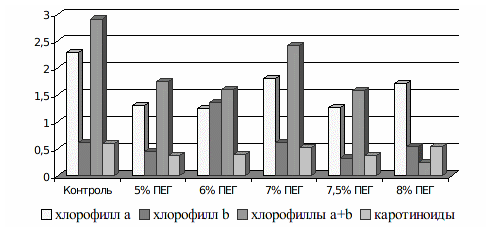
Рисунок 3. - Содержание пигментов в тканях листьев микроклонов винограда сорта Добрыня под влиянием модельной засухи, мг/г сырого веса.
Таблица 2 - ВЛИЯНИЕ МОДЕЛЬНОЙ ЗАСУХИ НА ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ И СОДЕРЖАНИЕ ВОДЫ В ЛИСТЬЯХ МИКРОКЛОНОВ ВИНОГРАДА
|
Вариант |
Интенсивность транспирации за 0,5 час, г/м2 · час | Содержание легкоудерживаемой воды, % |
Содержание общей воды, % |
Добрыня | |||
|
Контроль 5,0% ПЭГ 6,0% ПЭГ 7,0% ПЭГ 7,5% ПЭГ 8,0% ПЭГ |
62,061 72,656 52,620 86,092 58,923 130,400 |
63,5 38,6 41,3 43,2 32,1 43,6 |
87,6 75,2 79,7 77,6 76,1 71,9 |
Таировский 1 | |||
|
Контроль 5,0% ПЭГ 6,0% ПЭГ 7,0% ПЭГ 7,5% ПЭГ 8,0% ПЭГ |
41,760 42,064 56,461 45,759 56,849 67,101 |
52,1 36,2 34,2 36,1 44,5 37,9 |
84,9 80,2 72,3 75,7 78,3 71,2 |
Таким образом, проведенные исследования показывают возможность применения методов ИФХ листьев и создания условий водного стресса с использованием ПЭГ различных концентраций для изучения влияния почвенной засухи и определения реакции растений винограда на ее действие. Методы можно успешно применять для исследований пластичности сорта в изменяющихся условиях окружающей среды.
Литература
-
Брайон О.В., Корнеєв Д.Ю., Снегур О.О., Китаєв О.І. Інструментальне вивчення фотосинтетичного аппарату за допомогою індукції флуоресценції хлорофілу. // Методичні вказівки для студентів біологічного факультету. – К.: Видавничо-поліграфічний центр «Київський університет», 2000. – 15 с.
-
Григорюк И.А., Ткачев В.И. и др. Методы исследования и способы оценки устойчивости растений к засухе и высокой температуре. – К.: Знання, 1999. – 90 с.
-
Кондо И.Н. Устойчивость виноградного растения к морозам, засухе и почвенному засолению. – Кишинев: Картя Молдовеняске, 1970. – 96 с.
-
Косаківська І.В. Фізіолого-біохімічні основи адаптації рослин до стресів. – Київ: Сталь, 2003. – 191 с.
-
Рубин А.Б. Первичные процессы фотосинтеза // Соровский образовательный журнал. – 1997. - №10. – с. 79-84.
-
Савельев Н.И., Кузнєцова Н.В. Устойчивость семечковых культур к засухе и возможности ее повышения //Садоводство и виноградарство. – 2008. - №3. – С. 11-13.
-
Третьяков И.Н., Карнаухов Т.В., Паничкин Л.А. и др. Практикум по физиологии растений. – М.: Агропромиздат, 1990.
-
Физиология винограда и основы его возделывания. / Под рук. и ред. Стоева К. - Т.3 – София: Изд-во Болгарской академии наук, 1984. – 328 с.
- Войдите или зарегистрируйтесь, чтобы получить возможность отправлять комментарии
- 2398 reads
Последние новости
- АЗОСВиВ РЕАЛИЗУЕТ САЖЕНЦЫ!
- Свидетельство программы для ЭВМ "ПРОГНОЗИРОВАНИЕ УРОЖАЙНОСТИ СОРТОВ ВИНОГРАДА"
- 28 февраля состоялось совещание посвященное развитию виноградарско-винодельческой отрасли на территории муниципального образования г-к Анапа
- Авторское свидетельство на программу для ЭВМ "Автоматизированная система кодификации сортов винограда"
- Авторское свидетельство на программу для ЭВМ «АВТОМАТИЗИРОВАННАЯ СИСТЕМА СБОРА, УЧЕТА И ОБРАБОТКИ ДАННЫХ АГРОБИОЛОГИЧЕСКИХ И ФЕНОЛОГИЧЕСКИХ ПОКАЗАТЕЛЕЙ СОРТОВ ВИНОГРАДА»
- Красностоп Анапский Вести. Кубань
- 40-й Международный конкурс вин "Ялта золотой грифон 2020"
- Информация о переносе и изменении формата проведения международной научной конференции, посвященной 110-летию со дня образования АЗОСВиВ - филиала ФГБНУ СКФНЦСВВ
- МЕЖДУНАРОДНАЯ НАУЧНАЯ КОНФЕРЕНЦИЯ «ПОВЫШЕНИЕ ЭФФЕКТИВНОСТИ ИСПОЛЬЗОВАНИЯ БИОРЕСУРСОВ НА ОСНОВЕ БИОТЕХНОЛОГИЧЕСКИХ МЕТОДОВ СЕЛЕКЦИИ И РАЗМНОЖЕНИЯ ВИНОГРАДА»
- Сюжет на телеканале Кубань-24
Вход в систему
Разработка и поддержка сайта - АЗОС ВиВ